Calycadenia pauciflora
in W. H. Emory, Rep. U.S. Mex. Bound. 2(1): 100. 1859.
Plants 10–50 cm; self-incompatible. Stems branched (plants without obvious main axes, branches usually relatively many, slender, usually divergent, often zigzag), finely strigose (some hairs longer). Leaves mostly alternate, 1–5 cm (proximal), ± hispidulous and ± long-hairy (proximal margins and adaxial faces). Heads borne singly or in open, spiciform arrays (1–4 per node). Peduncular bracts ± linear to narrowly elliptic to oblanceolate, 2–3 mm (hispidulous, ± bristly and/or pectinate-fimbriate), apices rounded to truncate, tack-glands 1–3+. Phyllaries 4–6 mm, abaxial faces scabrous, ± strongly bristly, ± shaggy long-hairy on distal margins, tack-glands (0–) 1–7+. Paleae 4–6 mm (receptacular cups ± clavate to fusiform, lengths ± 2 times diams.). Ray-florets 1 (–2); corollas white to reddish, tubes ca. 2 mm (papillate), laminae 5–7 mm (central lobes equaling or slightly narrower than laterals, ± elliptic or distally broader and more rounded, symmetric or nearly so, widest near or beyond middles, sinuses nearly equaling laminae). Disc-florets 2–5; corollas white to pinkish, ca. 5–6 mm. Ray cypselae 2.5–4 mm, smooth, usually glabrous, sometimes hairy distally, sometimes with slightly elevated areolae. Disc cypselae angular, 2.5–3.5 mm, appressed hairy, especially on angles; pappi of ca. 8–10 lanceolate-attenuate and shorter, blunt fimbriate scales ca. 1–2.5 mm (alternating). 2n = 10, 12.
Phenology: Flowering spring–late summer.
Habitat: Open, dry, usually rocky meadows, hillsides
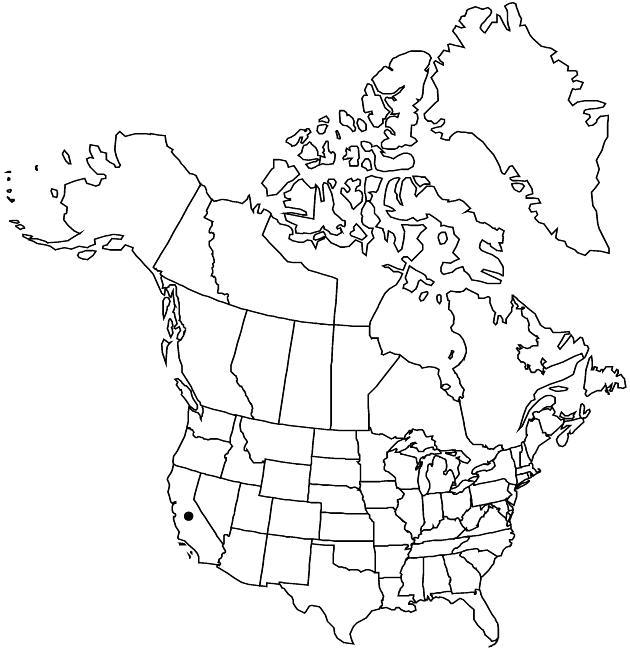
Elevation: 50–1000 m
Discussion
Calycadenia pauciflora is variable and closely related to and intergrades with C. fremontii. It is found mostly in the foothills of the Coast Ranges from Tehama County to Lake, Napa, and Yolo counties. It is also known from one location on the valley floor in Butte County.
The morphologic and chromosomal similarity of some populations of Calycadenia ciliosa to C. fremontii led R. L. Carr and G. D. Carr (1983) to merge the two species. Traditional treatments of these taxa and C. pauciflora have placed together populations that are far from homogeneous and may vary from one another in morphology and chromosome number and structure. For instance, C. fremontii and C. pauciflora are each known to include at least six different chromosome races. In this treatment, we have chosen to recognize the larger-headed members of C. pauciflora in the sense of G. D. Carr (1975) as C. fremontii. This reflects the current state of our knowledge, including molecular studies (B. G. Baldwin and S. Markos 1998). Molecular studies have provided a separate means of assessing relationships and insights into the evolution of the genus. Providing a practical taxonomic framework that reflects what is now known about the complicated morphologic and chromosomal patterns of variation of C. pauciflora and C. fremontii has proved to be an elusive goal.
Selected References
None.