Cirsium altissimum
Syst. Veg. 3: 373. 1826.
Biennials or short-lived monocarpic perennials, (50–) 100–300 (–400) cm; taproots and often a cluster of coarse fibrous-roots, roots without tuberlike enlargements. Stems single, erect, villous with septate trichomes, sometimes ± glabrate, sometimes distally thinly tomentose; branches few–many, ascending. Leaves: blades oblanceolate to elliptic or ovate, 10–40 × 1–13 cm, margins flat, finely spiny-toothed and otherwise undivided to coarsely toothed or shallowly pinnatifid, lobes broadly triangular, main spines 1–5 mm, abaxial faces white-tomentose, adaxial faces green, glabrate to villous with septate trichomes; basal usually absent at flowering, winged-petiolate, bases tapered; principal cauline well distributed, gradually reduced, bases narrowed, sometimes weakly clasping; distal cauline well developed. Heads 1–many, in corymbiform or paniculiform arrays, (± elevated above principal cauline leaves), not subtended by ring of spiny bracts. Peduncles 0–5 cm (leafy-bracted). Involucres ovoid to broadly cylindric or campanulate, (2–) 2.5–3.5 (–4) × (1.5–) 2–3 (–4) cm, thinly arachnoid. Phyllaries in 10–20 series, strongly imbricate, greenish with subapical darker central zone, ovate (outer) to lanceolate (inner), abaxial faces with a narrow glutinous ridge (milky when fresh, dark when dry), outer and middle entire, bodies appressed, spines slender, abruptly spreading, 3–4 mm; apices of inner phyllaries spreading, narrow, flattened, entire, spines spreading, slender, 3–4 mm; apices of inner phyllaries spreading, narrow, flattened, ± dilated, ± erose or finely serrulate. Corollas pink to purple (white), 20–35 mm, tubes 10–16 mm, throats 5–12 mm, lobes 5–9 mm. Style tips 4–6 mm. Cypselae tan to dark-brown, 4–5.5 mm, apical collars stramineous, 0.5–1 mm; pappi 12–24 mm. 2n = 18.
Phenology: Flowering summer–fall (Jun–Oct).
Habitat: Prairies, woodlands, disturbed sites, often in damp soil
Elevation: 50–700 m
Distribution
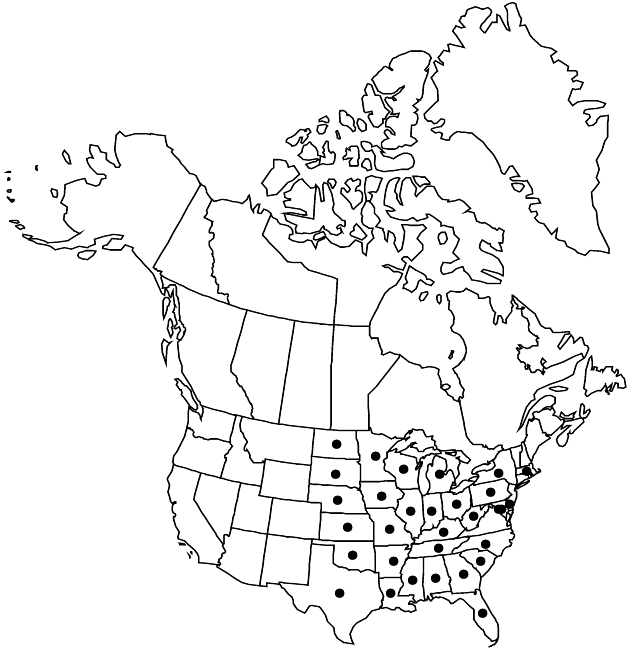
Ala., Ark., Del., D.C., Fla., Ga., Ill., Ind., Iowa, Kans., Ky., La., Md., Mass., Mich., Minn., Miss., Mo., Nebr., N.Y., N.C., N.Dak., Ohio, Okla., Pa., S.C., S.Dak., Tenn., Tex., W.Va., Wis.
Discussion
Plants of Cirsium altissimum ranging from southern Minnesota to Texas often have more deeply divided leaves than do populations in other portions of the species’ range. Some botanists (e.g., R. J. Moore and C. Frankton 1969; D. S. Correll and M. C. Johnston 1970) have treated those plants as C. iowense. Others (e.g., R. E. Brooks 1986; H. A. Gleason and A. Cronquist 1991; G. B. Ownbey and T. Morley 1991) have treated them as C. altissimum. Still others considered them to be derivatives of hybridization between C. altissimum and C. discolor (J. T. Kartesz and C. A. Meacham 1999) and treated them as C. ×iowense. Indeed the existence of these plants blurs the distinction between C. altissimum and C. discolor, and herbarium specimens are often difficult to assign.
Natural hybrids between Cirsium altissimum and C. discolor are well documented (R. A. Davidson 1963; G. B. Ownbey and Hsi Y.-T. 1963; Ownbey 1964; S. Dabydeen 1997). Ownbey and Dabydeen both reported that apparent F1 hybrids between the two species have low seed set in comparison with the parental taxa. W. L. Bloom (1977) reported that the chromosomes of the two species differ by several rearareaments. Dabydeen reported a count of 2n = 19 with multiple meiotic irregularities for an apparent F1 hybrid. However, the presence of numerous individuals and populations seemingly intermediate between C. altissimum and C. discolor indicates that although F1 hybrids have low fertility, long-term processes may have stabilized hybrid derivatives of higher fertility. Ownbey and Hsi reported mitotic counts of 2n = 18 and 20 from a population that they treated as C. altissimum. In their discussion they noted that their plants represented “the segregate called C. iowense” and had been collected a short distance from that taxon’s type locality. R. J. Moore and C. Frankton (1969) reported a chromosome number of 2n = 18 for a plant from Texas that they considered to be C. iowense. Further investigation of morphologic variation, chromosome number, meiotic behavior, and fertility is needed of populations named as C. iowense to determine how those plants should be treated.
Selected References
None.
Lower Taxa
"fine" is not a number.