Salix silicicola
J. Arnold Arbor. 17: 236, plate 194. 1936.
Shrubs, 1–3 m, (forming clones by layering). Stems: branches yellowbrown, gray-brown, or redbrown, not glaucous, villous in patches to glabrescent; branchlets gray-brown or redbrown, very densely villous. Leaves: stipules foliaceous, apex acute to acuminate; petiole convex to flat adaxially, 5–12 mm, villous or tomentose adaxially, (strongly ventricose around floral buds); largest medial blade (apparently hypostomatous but surfaces obscured by hairs), narrowly oblong, narrowly elliptic, elliptic, or obovate, 36–84 × 19–40 mm, 1.8–3.6 times as long as wide, base convex or cuneate, margins slightly revolute, entire, apex convex or acuminate, abaxial surface very densely woolly-tomentose, hairs wavy, adaxial dull, moderately to very densely, villous-tomentose; proximal blade margins entire; juvenile blade color obscured by hairs, very densely tomentose-woolly abaxially, hairs white. Catkins flowering before leaves emerge; staminate stout, 40–56 × 14–15 mm, flowering branchlet 0 mm; pistillate densely flowered, slender, 35–125 (–130 in fruit) mm, flowering branchlet 0 mm; floral bract brown or black, 2–3 mm, apex convex to rounded, abaxially hairy, hairs straight. Staminate flowers: adaxial nectary oblong to narrowly oblong, 0.6–1.5 mm; filaments distinct; anthers yellow, ellipsoid, 0.6–0.7 mm. Pistillate flowers: adaxial nectary oblong or obtriangular, 0.5–1.1 mm; stipe 0–0.3 mm; ovary pyriform, beak gradually tapering to styles; ovules 12–14 per ovary; styles 1.2–2.2 mm; stigmas 0.52–0.75–1 mm. Capsules 4–7 mm. 2n = 38.
Phenology: No data are available on flowering time in the wild; in cultivation flowering is early May.
Habitat: Active sand dunes
Elevation: 20-500 m
Discussion
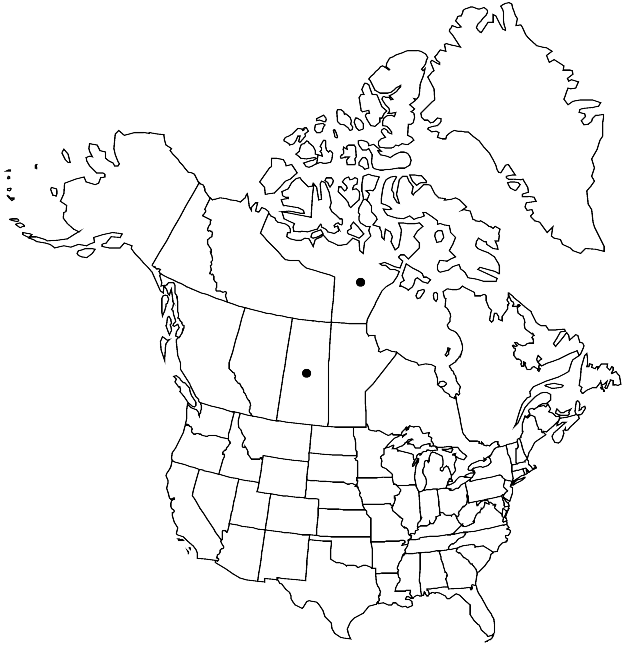
Comparisons of genetic variation in Salix alaxensis var. alaxensis from British Columbia and S. silicicola from Lake Athabasca sand dunes, Saskatchewan, based on isozyme loci, fit predicted relationships between progenitor and derived taxon (B. G. Purdy and R. J. Bayer 1995). Allelic diversity of S. silicicola was a subset of that of S. alaxensis, there was less genetic variation in S. silicicola than in S. alaxensis, and interspecific genetic variation within the two species was similar and relatively very high. This suggested a recent origin for the derived S. silicicola.
Salix silicicola is a uniform population that differs from S. alaxensis in its very densely villous or tomentose leaves and branchlets. These characters seem to be an adaptation to reduce sand abrasion and water loss in a sand dune environment. It is unlikely that it would have evolved in situ but probably derived from a pre-adapted source such as the one represented by specimens of putative S. silicicola from Pelly Lake, Nunavut. The isozyme study did not include specimens from that population or of S. alaxensis from Northwest Territories from which S. silicicola is likely to have been derived. Occurrence of S. silicicola-like plants in northern continental Nunavut suggests that during the late Pleistocene, it had a wider range, which now is represented by two disjunct populations. The question of appropriate taxonomic rank for the derived taxon is still unresolved. Although S. silicicola is different from S. alaxensis in its general appearance, they are very similar genetically, and argument could be made for treating them as varieties (B. Boivin 1966b).
Hybrids:
Salix silicicola forms natural hybrids with S. brachycarpa var. psammophila.
of conservation concern
Selected References
None.
Lower Taxa
"-1mm" is not declared as a valid unit of measurement for this property.