Carex diluta
Fl. Taur.-Caucas. 2: 388. 1808.

Culms 15–55 cm. Leaves mostly basal; basal sheaths pale to dark-brown; blades of flowering-stems yellowish green, flat, shorter than stems, widest blades 3–4.6 mm wide, margins scabrous. Inflorescences: longer peduncle of terminal spike 1–2 (–4) cm; proximal bract mostly equaling or no more than 1.5 times as long as inflorescence; pistillate spikes (2–) 3–5, ascending, ovoid to short-cylindric; distal 1 or 2 pistillate spikes aggregated near staminate spike; the proximal widely separated, pedunculate; terminal staminate spike 8–25 (–30) × 2 mm. Pistillate scales brown to reddish-brown, with green or whitish midrib. Staminate scales brown, ovate-elliptic, margins scarious, apex obtuse or mucronate. Anthers 1.5–2.7 mm. Perigynia yellowish green, 2.6–3.5 × 1–1.3 mm, apex gradually tapered; beak 0.4–0.7 mm, smooth.
Phenology: Fruiting summer (Aug–early Sep).
Habitat: Open, damp, saline meadows and bottomlands
Elevation: 1300–1400 m
Distribution
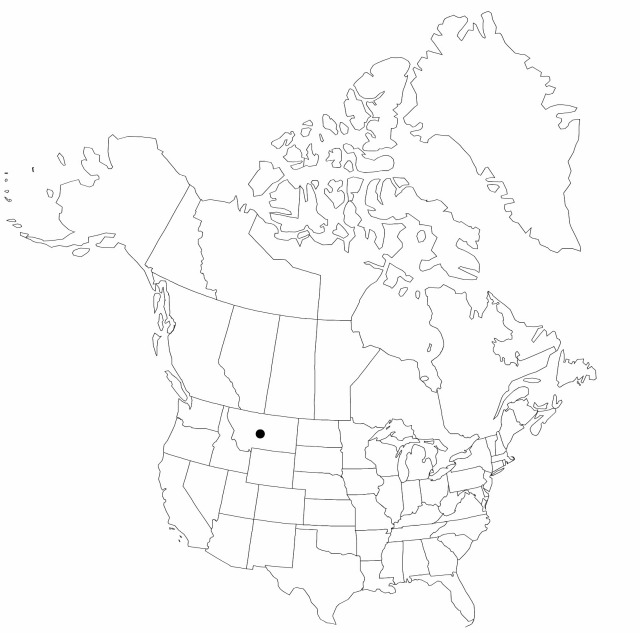
Introduced; Mont., Eurasia
Discussion
Carex diluta was discovered in Montana in 1995; it occurs there in relatively natural saline habitats. Its similarity to some Eurasian members of the C. diluta complex suggests that it is probably an introduction.
The taxonomy of Carex diluta and closely related taxa varies throughout Eurasia. A. O. Chater (1980) and T. V. Egorova (1999) applied a broad species concept, which is followed here. However, V. I. Kreczetowicz (1935) recognized four taxa in the complex: C. diluta (in the strict sense), C. karelinii Meinshausen, C. aspratilis V. I. Kreczetowicz, and C. czarwakensis Litvinov. Montana populations appear to correspond most closely to C. karelinii. Further comparisons of the Montana plants with Eurasian material are needed, and indeed, a further examination of this complex throughout its native range in Eurasia is warranted.
Selected References
None.
Lower Taxa
"shortened" is not a number."+timesaslongasinflorescence" is not declared as a valid unit of measurement for this property.