Eleocharis kamtschatica
Fl. Kamtschatka 1: 207. 1927.

Plants perennial, mat-forming; rhizomes evident, sometimes vertical, long, 0.5–1.5 mm thick, soft, cortex often breaking loose, longer internodes 1–3 cm, scales fugaceous, 4–7 mm, thinly membranous, not fibrous. Culms terete, often with to 10 blunt ridges when dry, 4–60 cm × 0.3–1.8 mm, soft to firm, internally spongy. Leaves: distal leaf-sheaths persistent, not splitting, proximally red (to stramineous), distally red or stramineous or green, not callose, membranous, apex usually pale-red, broadly obtuse to narrowly acuminate, tooth absent. Spikelets ovoid to lanceoloid, 4–20 × 3–6 mm, apex acute; proximal scale amplexicaulous, entire; subproximal scale with flower; floral scales often spreading in fruit, 5–20, 1–2 per mm of rachilla, entirely blackish brown to redbrown, or midrib regions paler, broadly ovate, 2.5–5 × 2–2.5 mm, apex rounded to subacute, not carinate. Flowers: perianth bristles 0–6 (–9), light-brown to stramineous, slender to stout, often unequal, much shorter than to equaling achene; stamens 3; anthers yellowbrown to medium brown, 1.3–2.5 mm; styles 2-fid. Achenes not persistent, yellow, stramineous, or medium brown, obovoid (mostly very broadly so), thickly biconvex, angles obscure, 1–1.5 (–1.9) × 1–1.6 mm, apex truncate, neck absent or very short, finely reticulate at 20–30X, or rugulose with 20 or more horizontal ridges in a vertical series, or sometimes smooth. Tubercles whitish, outline subrectangular or trapezoidal, sometimes broadly rounded, seldom pyramidal, (0.5–) 0.7–1.5 × (0.5–) 0.8–1.4 mm, usually with vertical rows of depressions (cancellate). 2n = 12.
Phenology: Fruiting summer.
Habitat: Brackish (to fresh?) marshes, meadows, ponds, inland in Asia
Elevation: 0–30 m
Distribution
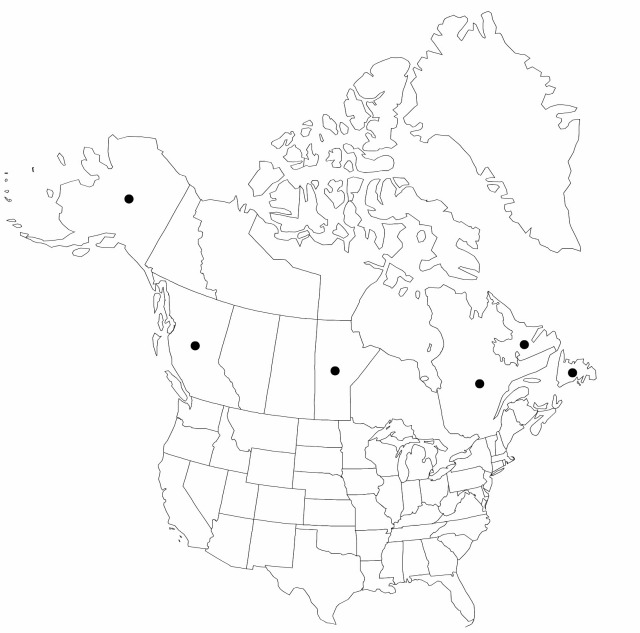
B.C., Man., Nfld. and Labr., Que., Alaska, Asia (China), Asia (Japan), Asia (Korea), Asia (Russia)
Discussion
The markedly different chromosome counts reported from North America (2n = 12, from the Queen Charlotte Islands, British Columbia) and from Asia (2n = 38–40, 42, 56) suggest that more than one taxon should be recognized within Eleocharis kamtschatica as currently broadly defined. The three similar species E. kamtschatica, E. komarovii, and E. sachalinensis have been recognized for Russia (I. D. Zinserling 1935); more recent authors have recognized only E. kamtschatica (T. V. Egorova 1981 and A. E. Kozhevnikov 1988, for the former U.S.S.R.; J. Ohwi 1965, for Japan; S.-O Strandhede 1967; E. Hultén 1968 for Alaska; Tang T. and Wang F. T. 1961, for China). Pending a taxonomic revision, the following five names, all from eastern Asia, should probably be treated as synonyms of E. kamtschatica: E. komarovii Zinserling, E. mitrata (Franchet & Savatier ex Makino, E. sachalinensis (Meinschauser) B. Fedtschenko, E. savatieri Svenson, and E. triflora Komarov, not E. triflora Boeckeler. In E. kamtschatica in the broad sense, the size and form of the achenes and tubercles, on which most of the probable synonyms listed above are based, and the form of the distal leaf sheath summits do not seem to be correlated with each other or with other characters. In North America E. kamtschatica is known only from coastal habitats; in Asia it also occurs inland, especially around hot springs.
North American plants of Eleocharis kamtschatica seem to be separable into two groups: (1) Smaller plants, with culms mostly shorter than 20 cm, spikelets 5–13 × 3–4 mm, and floral scales 2.5–4 mm are known from about 20 localities on the Pacific Coast from Kodiak Island, Alaska, to the Queen Charlotte Islands and Prince Rupert, British Columbia, as well as from Hudson Bay in Manitoba, James Bay and the Gulf of St. Lawrence in Quebec, and Labrador. These small plants include the voucher specimens of the 2n = 12 chromosome count from British Columbia, and seem to fit E. kamtschatica as described by I. D. Zinserling (1935). (2) More robust plants, with culms mostly 20–70 cm, spikelets 7–20 × 4–6 mm, and floral scales 4–5 mm, are known from about 12 localities from Norton Sound in west-central Alaska to central British Columbia. These robust plants fit the descriptions of E. sachalinensis by I. D. Zinserling(1935), of E. savatieri Svenson from Japan (H. K. Svenson 1939), and of E. kamtschatica as described by J. Ohwi (1965), and are very similar to several specimens from Japan that I have seen.
Because Eleocharis kamtschatica closely resembles E. uniglumis except for size of the tubercles, plants without achenes cannot be reliably identified to species. The floral scales and receptacles in E. kamtschatica are very sparsely placed, with only 1–2 scales per mm of rachilla, compared with 3–4 mm of rachilla in most North American plants of E. uniglumis. Some North American collections, e.g., from the Kobuk sand dunes in northwestern Alaska, as E. uniglumis, and James Bay in Quebec, as E. kamtschatica, are intermediate between E. kamtschatica and E. uniglumis in the relative sizes of the tubercles and achenes.
Selected References
None.
Lower Taxa
"shortened" is not a number.